Post by sebastian on Oct 24, 2006 3:12:56 GMT
Apteribis brevis, new species
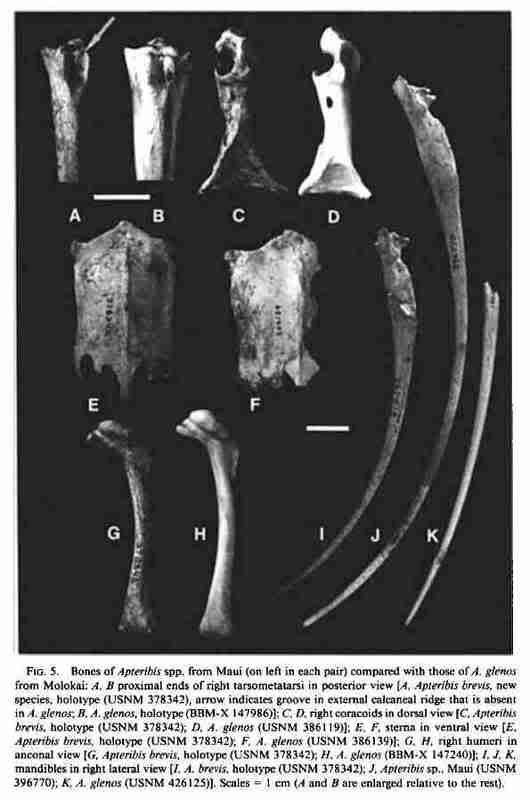
(Fig. 5A, C, E, G, I)
Apteribis glenos (part) Olson and Wetmore, 1976.
"Apteribis sp." Olson and James, 1982b:33; James et al., 1987:2353.
"Maui Apteribis sp. A" Olson and James, 1984: 772.
Holotype: Nearly complete associated skeleton, USNM 378342 (Fig. 5A, C, E, G, I). Collected 8 September 1982 by Storrs L. Olson and Helen F. James. The specimen includes the skull (lacking right quadrate, pterygoids, and most of both quadratojugals), mandible, notarium, pelvis, all major elements of the wing, pectoral girdle and hindlimb (except right femur), various vertebrae, ribs, and phalanges of manus and pes.
Type locality: Auwahi Cave (1,145 m), Maui, Hawaiian Islands.
Distribution: Maui: from higher elevations on the southern slopes of Mr. Haleakala, including upper Kipahulu and possibly Lower Waihoi Valleys.
Etymology: Latin, brevis, short, from the short bill and small size that characterize this species.
Measurements (mm) ofholotype: Skull: total length, 135.4; length of rostrum along midline from "forehead," 105.6; length and width of cranium, 36.5 x 31.6. Mandible: total length, 111.6; length of symphysis, 62.1; width of articulation, 10.9. Sternum: length along midline, 50.3; width through second costal facet, 29.2. Notarium: length ofcentra along midline, 26.4. Pelvis: length of synsacrum, 51.5; width through antitrochanters, 35.7. Coracoid: length, 29.8. Scapula: length, 42.2; width ofarticular end, 8.6. Furcula: length, 29.0. Humerus: length, 62.5; proximal width, 15.1; shaft width at midpoint, 5.0; distal width, 11.5. Ulna: length, 48.6; proximal depth, 8.5. Radius: length, 46.7. Carpometacarpus: length, 24.6; proximal depth, 8.5. Femur: length, 62.7; proximal width, 14.3; shaft width at midpoint, 6.1; distal width, 13.9. Tibiotarsus: length from cnemial crest, 94.0; shaft width at midpoint, 6.0; distal width, 11.2. Tarsometatarsus: length, 53.0; proximal width, 13.2; shaft width at midpoint, 6.8; distal width, 13.6.
Paratypes: A multitude of bones from many different individuals; virtually theentire skeleton is represented. The paratypes are not listed individually for economy of space. The incomplete associated skeleton from Lower Waihoi Valley Cave (BBM-X 147127-147175; see Olson and Wetmore 1976:248) is tentatively referred to this species based on size alone.
Measurements of paratypes: Certain measurements of long bones are incorporated into Tables 2 and 3 and Figures 5 and 6.
Diagnosis: Differs from Apteribis glenos as follows: tip of premaxilla and mandible not as narrow and pointed, mandible noticeably more decurved (Fig. 5I, K); sternum much more arched (concave dorsally), not broad and flattened as in A. glenos (Fig. 5E, F); coracoid with procoracoid foramen larger and situated closer to the medial margin of the bone, impression of M. sternocoracoideus extending farther up the shaft, not marked with a distinct semicircular lip, and decidedly more flattened at the sternal end, as opposed to concave (Fig. 5C, D); humerus (Fig. 5G, H) and ulna not as robust, humerus with head and pectoral crest less in flated; calcanear lidges of tarsometatarsus more elongate, the lateral one invariably with a distinct tendinal groove that is absent in A. glenos (Fig. 5A, B).
Remarks: Ibises were probably comparatively abundant on Maui, where they were undoubtedly birds of the forest floor. Being flightless, they were highly susceptible to being trapped in lava tubes. Ibises (e.g., Threskiornis aethiopicus, pers. observ.) may at times feed heavily on snails. Apteribis, along with rails, may thus have exerted considerable predation pressure on the endemic land snails of Maui Nui, a factor that malacologists should recognize when assessing evolution in Hawaiian pulmonates. Unfortunately, the land snails of the Hawaiian Islands experienced much the same fate that befell so many of the birds and it may never be known if the snails of Maui Nui had evolved behavioral traits different from those on islands where ibises were absent.
(Fig. 5A, C, E, G, I)
Apteribis glenos (part) Olson and Wetmore, 1976.
"Apteribis sp." Olson and James, 1982b:33; James et al., 1987:2353.
"Maui Apteribis sp. A" Olson and James, 1984: 772.
Holotype: Nearly complete associated skeleton, USNM 378342 (Fig. 5A, C, E, G, I). Collected 8 September 1982 by Storrs L. Olson and Helen F. James. The specimen includes the skull (lacking right quadrate, pterygoids, and most of both quadratojugals), mandible, notarium, pelvis, all major elements of the wing, pectoral girdle and hindlimb (except right femur), various vertebrae, ribs, and phalanges of manus and pes.
Type locality: Auwahi Cave (1,145 m), Maui, Hawaiian Islands.
Distribution: Maui: from higher elevations on the southern slopes of Mr. Haleakala, including upper Kipahulu and possibly Lower Waihoi Valleys.
Etymology: Latin, brevis, short, from the short bill and small size that characterize this species.
Measurements (mm) ofholotype: Skull: total length, 135.4; length of rostrum along midline from "forehead," 105.6; length and width of cranium, 36.5 x 31.6. Mandible: total length, 111.6; length of symphysis, 62.1; width of articulation, 10.9. Sternum: length along midline, 50.3; width through second costal facet, 29.2. Notarium: length ofcentra along midline, 26.4. Pelvis: length of synsacrum, 51.5; width through antitrochanters, 35.7. Coracoid: length, 29.8. Scapula: length, 42.2; width ofarticular end, 8.6. Furcula: length, 29.0. Humerus: length, 62.5; proximal width, 15.1; shaft width at midpoint, 5.0; distal width, 11.5. Ulna: length, 48.6; proximal depth, 8.5. Radius: length, 46.7. Carpometacarpus: length, 24.6; proximal depth, 8.5. Femur: length, 62.7; proximal width, 14.3; shaft width at midpoint, 6.1; distal width, 13.9. Tibiotarsus: length from cnemial crest, 94.0; shaft width at midpoint, 6.0; distal width, 11.2. Tarsometatarsus: length, 53.0; proximal width, 13.2; shaft width at midpoint, 6.8; distal width, 13.6.
Paratypes: A multitude of bones from many different individuals; virtually theentire skeleton is represented. The paratypes are not listed individually for economy of space. The incomplete associated skeleton from Lower Waihoi Valley Cave (BBM-X 147127-147175; see Olson and Wetmore 1976:248) is tentatively referred to this species based on size alone.
Measurements of paratypes: Certain measurements of long bones are incorporated into Tables 2 and 3 and Figures 5 and 6.
Diagnosis: Differs from Apteribis glenos as follows: tip of premaxilla and mandible not as narrow and pointed, mandible noticeably more decurved (Fig. 5I, K); sternum much more arched (concave dorsally), not broad and flattened as in A. glenos (Fig. 5E, F); coracoid with procoracoid foramen larger and situated closer to the medial margin of the bone, impression of M. sternocoracoideus extending farther up the shaft, not marked with a distinct semicircular lip, and decidedly more flattened at the sternal end, as opposed to concave (Fig. 5C, D); humerus (Fig. 5G, H) and ulna not as robust, humerus with head and pectoral crest less in flated; calcanear lidges of tarsometatarsus more elongate, the lateral one invariably with a distinct tendinal groove that is absent in A. glenos (Fig. 5A, B).
Remarks: Ibises were probably comparatively abundant on Maui, where they were undoubtedly birds of the forest floor. Being flightless, they were highly susceptible to being trapped in lava tubes. Ibises (e.g., Threskiornis aethiopicus, pers. observ.) may at times feed heavily on snails. Apteribis, along with rails, may thus have exerted considerable predation pressure on the endemic land snails of Maui Nui, a factor that malacologists should recognize when assessing evolution in Hawaiian pulmonates. Unfortunately, the land snails of the Hawaiian Islands experienced much the same fate that befell so many of the birds and it may never be known if the snails of Maui Nui had evolved behavioral traits different from those on islands where ibises were absent.