DISCUSSION
Flightlessness
Rails are developmentally predisposed to become flightless (Olson 1973b, Feduccia 2000), and scores of neotenic flightless species have evolved independently on oceanic islands that lack indigenous placcntal carnivores. The hypothesis that flightlessness evolves as a means of energy conservation is supported by the basal metabolic rates in flightless species of insular rails being lower than those of their volant relatives (McNab 1994a,b). This finding, coupled with the observation that metabolic rate also correlates positively with relative pectoral muscle mass (McNab 1994a, 2002), suggests that selection for reduced pectoral and wing musculature is the likely cause of insular avian flightlessness.
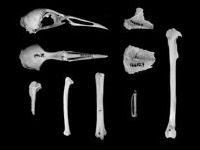
Skeletons of flightless rails are distinguished from those of volant relatives by having reduced sternal keels (carina sterni) and shorter, thinner wing bones. Livezey (1998, 2003) has shown that the degree of reduction of pectoral elements varies greatly even among flightless species. Indeed there appears to be a continuum of wing reduction among flightless rails that mirrors the graded reduction in energy expenditure (McNab 2002). Our morphometric analysis of Gallirallus skeletons indicates that some species, such as G. australis, have greatly reducedcarina sterni and wing elements, whereas other extant species known to be flightless, such as G. owstoni, seem to be near the threshold of flight.
Biogeography
Of the 18 living and extinct species of Gallirallus sensu lato that have been named, all but two are flightless species endemic to Oceania on single islands or on multiple islands that were connected during the late Pleistocene period of lowered sea levels (Steadman 1987, in press, Diamond 1991, Mayr and Diamond 2001). The two extant, volant species, G. torquatus and G. philippensis, are sympatric in the Philippines, Sulawesi, and New Guinea, although the latter species is very widespread, its distribution extending south to Australia and New Zealand and east in Oceania to Samoa. Flightless species of Gallirallus have evolved on nearly all major archipelagos in Oceania from the Ryukyu Islands of southern Japan, south to New Zealand's Chatham Islands, and east to the Society Islands. Bones (still undescribed) of Gallirallus also have been found in archaeological sites on four islands in the Marquesas (Steadman 1989a, Steadman and Rolett 1996). Gallirallus apparently never made it as far northeast in Oceania as the Hawaiian Islands or as far soudieast in Polynesia as Henderson Island, botii of which have good Holocene fossil bird records (James and Olson 1991, Olson and James 1991, Wragg 1995, Steadman in press).
Gallirallus stomolsoni is not the only species of flightless rail known from the Society Islands. The extinct G. pacificus was discovered on Tahiti by naturalists from Captain James Cook's second voyage (1777), but no specimens exist, and the species is known only from a painting by Georg Forster. The plumage, soft-part colors, and bill shape make it clear that pacificus is correctly accomodated in Gallirallus. Measurements made by Storrs L. Olson in 1998 (pers. comm.) from Forster's original, full-scale painting in the British Museum of Natural History indicate that G. pacificus was a much smaller rail than G. storrsolsoni. For example, the culmen (with epidermal sheath) in G. pacificus is 28.8 mm, whereas the rostrum (without epidermal sheath) in G. storrsolsoni is 36.5-41.0 mm (Table 1). The tarsus (incuding scutes) in G. pacificus is 33.8 mm, whereas the tarsometatarsus (without epidermal sheath) in G. storrsolsoni is ca. 48.5 mm, based on a composite of two incomplete specimens (DAPT 7, BPBM 166034). Gallirallus pacificus is presumed to be flightless on the basis of the short wings in the painting; although this is likely, in the absence of specimens it cannot be verified in the painting. The possible former existence of G. pacificus on Mehetia (Taylor 1998), a small island 110 km east-southeast of Tahiti, is unsubstantiated and doubtful.
Based on its geological and geographical setting, any flightless species on Huahine is likely to have been endemic to the island. The geological age of Huahine is ca. 3 million yr (Dickinson 1998). It is an eroded volcanic island surrounded by a broad fringing reef, outside of which the water becomes very deep. The nearest island is Ra'iatea, ca. 35 km to the west. Huahine never was connected to any other island, even during the lowered sea levels of Pleistocene glacial intervals, when Ra'iatea still would have been ca. 30 km away.
The genus Porphyrio is widespread in tropical and subtropical lowlands. Two extant, volant species, P. martinicus and P. flavirostris, are confined to the Americas. All other species are from the Old World, with extant, volant P. alleni in Africa and the larger, extant, volant P. porphyrio very widespread from southern Europe and Africa eastward across southern Asia, Indonesia, and Australia to Oceania as far east as Fiji, Tonga, Samoa, and Niue (Taylor 1998:464, 465). No species of Porphyrio inhabit East Polynesia today. The radiation of certainly or presumably flightless species of Porphyrio is confined to Oceania (Balouet and Olson 1989, Worthy and Holdaway 2002, Steadman in press). The only flightless species that still exists is P. hochstetteri from South Island, New Zealand. Known extinct forms of Porphyrio are P. mantdli (North Island, New Zealand), P. albus (Lord Howe Island), P. kukwiedii (New Caledonia), and Porphyrio undescribed sp. A (New Ireland) and sp. B (Buka, Solomon Islands). Each of these species was as large as or larger than the massive P. hochstetteri, which is the largest extant form.
Finally, Porphyrio paepae was a smaller, probably flightless swamphen that is known from Hiva Oa and Tahuata, two islands only 3 km from each other in the Marquesas (Steadman 1988). At the time of its description, P. paepae was the only species of Porphyrio known from East Polynesia. The discovery of P. mcnabi in the Society Islands helps to bridge the formerly huge distributional gap of the genus (from Niue to the Marquesas) and strengthens the likelihood that a substantial radiation of swamphens once existed in East Polynesia. At this point, we cannot say whether this radiation was of Old World or New World origin. Osteological synapomorphies that ally P. paepae and P. mcnabi with either the New World species (P. martinicus, P. flavirostris) or the Old World P. porphyrio s.l. have not been discerned. Given the propensity in P. martinicus and P. flavirostris for unpredictable, long-distance dispersal (Remsen and Parker 1990), the geographic origin of East Polynesian Porphyrio species remains an open question.
Avian Extinction on Huahine
The Fa'ahia archaeological site contained the bones of 15 resident species of seabirds, three migratory species of shorebirds, and 16 species of resident land birds (Steadman and Pahlavan 1992, Steadman in press). Of the 15 seabirds, 12 no longer occur on Huahine, including the extinct gull Larus utunui (Steadman 2002). The other 11 species of seabirds still are found, at least locally, elsewhere in Oceania. Each of the migratory shorebirds, on the other hand, may visit Huahine seasonally today. Of the 16 species of land birds from Fa'ahia, 13 no longer occur on Huahine. They consist of the locally extripated heron Butorides (Ardeola) striants, rail Porzana tabuensis, dove Gallicolumba erythroptera, pigeons Ducula galeata and D. aurorae, and warbler Acrocephalus caffer. Aside from Gallirallus storrsokoni and Porphyrio mcnabi, the extinct species of land birds from Fa'ahia include the doves Gallicolumba nui and Macropygia arevarevauupa, lorikeets Vini vidivici and V. sinotoi, and starling Aplonis diluvialis (Steadman and Zarriello 1987, Steadman 19896, 1992).
The survivors are the very widespread heron Egretta sacra and two species endemic to the Society Islands, the dove Ptilinopus purpuratus and kingfisher Halcyon tuta. Also extant on Huahine is the very widespread duck Anas superciliosa, which was not found among the bird bones from Fa'ahia. Two additional species of land birds, the lorikeet Vini peruviana and swiftlet Collocalia leucophaea, are unknown at the Fa'ahia site but were recorded from Huahine in the nineteenth century, although they no longer persist on the island (Steadman in press). Altogether, 19 species of land birds have been recorded from Huahine, which is the same number that is known from much-larger Tahiti, which has no fossil record of birds. If the prehistoric bird community of Huahine were entirely revealed, we believe that it also would include a species of Prosobonia sandpiper, Cyanoramphus parrot, and Pomarea monarch, three genera recorded on at least two other islands in the Society Group.
ACKNOWLEDGMENTS
For access to modern specimens, we thank the curatorial staffs of the eight museums listed in Materials and Methods. Carla Kishinami and Yosihiko Sinoto kindly provided access to the prehistoric bones from Huahine. We are grateful to Storrs Olson for sharing his measurements of Forster's painting of Gallirallus pacificus. We thank Storrs Olson and an anonymous reviewer for insightful comments that improved the manuscript.
1 Financial support came from a McGlaughlin Fellowship to J.J.K. from the University of Florida College of Liberal Arts and Sciences, and NSF grants EAR9714819 and DEB-0228682 to D.W.S. Manuscript accepted 20 June 2005.
Literature Cited
Balouet, J. C., and S. L. Olson. 1989. Fossil birds from late Quaternary deposits in New Caledonia. Smithson. Contrib. Zool. 469.
Baumel, J. L., and L. M. Witmer. 1993. Osteologia. Pages 45-132 in J. J. Baumel, A. S. King, J. E. Breazile, H. E. Evans, and J. C. Vanden Berge, eds. Handbook of avian anatomy: Nomina anatomica avium, 2nd ed. Nuttall Ornithological Club, Cambridge, Massachusetts.
Conte, E., and A. Anderson. 2003. Radiocarbon ages for two sites on Ua Huka, Marquesas. Asian Perspect. 42:155-160.
Diamond, J. M. 1991. A new species of rail from the Solomon Islands and convergent evolution of insular flightlessness. Auk 108:461-470.
Dickinson, W. R. 1998. Geomorphology and geodynamics of the Cook- Austral Island Seamount Chain in the South Pacific Ocean: Implications for hotspots and plumes. Int. Geol. Rev. 40:1039-1075.
Dye, T., and D. W. Steadman. 1990. Polynesian ancestors and their animal world. Am. Sci. 78:209-217.
Feduccia, A. 2000. The origin and evolution of birds, 2nd ed. Yale University Press, New Haven, Connecticut.
James, H. F., and S. E. Olson. 1991. Description of thirty-two new species of birds from the Hawaiian Islands: Part II. Passer- iformes. Ornithol. Monogr. 46.
Kirch, P. V. 1984. The evolution of the Polynesian chiefdoms. Cambridge University Press, Cambridge, United Kingdom.
_____. 1986. Rethinking East Polynesian prehistory. J. Polynesian Soc. 95:9-40.
_____. 2000. On the road of the winds: An archaeological history of the Pacific islands before European contact. University of California Press, Berkeley.
Kirch, P. V., and J. Ellison. 1994. Palaeoenvironmental evidence for human colonization of remote Oceanic islands. Antiquity 68:310- 321.
Kirch, P. V., D. W. Steadman, V. L. Butler, J. Hather,and M. Weisler. 1995. Prehistory and human ecology in eastern Polynesia: Excavations at Tangatatau Rockshelter, Mangaia, Cook Islands. Archaeol. Oceania 30:47-65.
Kirchman, J. J., and D. W. Steadman. 2005. Rails (Aves: Rallidae: Gallirallus) from prehistoric sites in the Kingdom of Tonga, including description of a new species. Proc. Biol. Soc. Wash. 118:465-477.
Lepofsky, D. 1995. A radiocarbon chronology for prehistoric agriculture in the Society Islands, French Polynesia. Radiocarbon 37:917-930.
Lepofsky, D., H. C. Harries, and M. Kellum. 1992. Early coconuts on Mo'orea Island, French Polynesia. J. Polynesian Soc. 101: 299- 308.
Lepofsky, D., P. V. Kirch, and K. P. Lertzman. 1996. Stratigraphie and paleobotanical evidence for prehistoric humaninduced environmental disturbance on Mo'orea, French Polynesia. Pac. Sci. 50: 253-273.
Livezey, B. C. 1998. A phylogenetic analysis of the Gruiformes (Aves) based on morphological characters, with an emphasis on the rails (Rallidae). Philos. Trans. R. Soc. Eond. B Biol. Sa. 353:2077- 2151.
_____. 2003. Evolution of Sightlessness in rails (Gruiformes: Rallidae): Phylogenetic, ecomorphological, and ontogenetic perspectives. Ornithol. Monogr. 53.
Mayr, E., and J. Diamond. 2001. The birds of northern Melanesia. Oxford University Press, Oxford, United Kingdom.
McNab, B. K. 1994a. Energy conservation and the evolution of Sightlessness in birds. Am. Nat. 144:628-642.
_____. 1994b. Resource use and the survival of land and freshwater vertebrates on oceanic islands. Am. Nat. 144:643-660.
_____. 2002. The physiological ecology of vertebrates: A view from energetics. Cornell University Press, Ithaca, New York.
Olson, S. L. 1973a. A classification of the Rallidae. Wilson Bull. 85:381-416.
_____. 1973b. Evolution of the rails of the South Atlantic islands (Aves: Rallidae). Smithson. Contrib. Zool. 152.
Olson, S. L., and H. F.James. 1991. Description of thirty-two new species of birds from the Hawaiian Islands: Part I. Non- Passeriformes. Ornithol. Monogr. 46.
Remsen, J. V., Jr., and T. A. Parker III. 1990. Seasonal distribution of the azure gallinule (Porphyrula flavirostris), with comments on vagrancy in rails and gallinules. Wilson Bull. 102:380- 399.
Sinoto, Y. H. 1970. An archaeologically based assessment of the Marquesas Islands as a dispersal center in East Polynesia. Pac. Anthropol. Rec. 11:105-132.
_____. 1975. Report on the preliminary excavation of an early habitation site on Huahine, Society Islands. J. Soc. Ocan. 31:143- 186.
_____. 1979. Excavations on Huahine, French Polynesia. Pac. Stud. 3:1-40.
_____. 1983. Archaeological excavations of the Vaito'tia and Fa'ahia sites on Huahine Island, French Polynesia. Nad. Geogr. Res. 15:583-599.
Spriggs, M., and A. Anderson. 1993. Late colonization of East Polynesia. Antiquity 67:200-217.
Steadman, D. W. 1987. Two new species of rails (Aves: Rallidae) from Mangaia, southern Cook Islands. Pac. Sci. 40:38-54.
_____. 1988. A new species of Porphyrio (Aves: Rallidae) from archaeological sites in the Marquesas Islands. Proc. Biol. Soc. Wash. 101:162-170.
_____. 1989a. Extinction of birds in eastern Polynesia: A review of the record, and comparisons with other Pacific island groups. J. Archaeol. Sci. 16:177-205.
______. 1989b. A new species of starling (Sturnidae, Aplanis) from an archaeological site on Huahine, Society Islands. Notornis 36:161-169.
_____. 1992. New species of Gallicolumba. and Macropygia (Aves: Columbidae) from archaeological sites in Polynesia. Los Angel. Cty. Mus. Nat. Hist., Sci. Ser. 36:329-348.
_____. 1995. Prehistoric extinctions of Pacific island birds: Biodiversity meets zooarchaeology. Science (Washington, D.C.) 267:1123-1131.
_____. 2002. A new species of gull (Laridae: Lams) from an archaeological site on Huahine, Society Islands. Proc. Biol. Soc. Wash. 115:1-17.
_____. In press. Extinction and biogeography of tropical Pacific birds. University of Chicago Press, Chicago.
Steadman, D. W., and D. S. Pahlavan. 1992. Extinction and biogeography of birds on Huahine, Society Islands, French Polynesia. Geoarchaeology 7:449-483.
Steadman, D. W., and B. Rolett. 1996. A chronostratigraphic analysis of landbird extinction on Tahuata, Marquesas Islands. J. Archaeol. Sci. 23:81-94.
Steadman, D. W., and M. C. Zarriello. 1987. Two new species of parrots (Aves: Psittacidae) from archaeological sites in the Marquesas Islands. Proc. Biol. Soc. Wash. 100:518-528.
Steadman, D. W., T. H. Worthy, A. J. Anderson, and R. Walter. 2000. New species and records of birds from prehistoric sites on Nine, Southwest Pacific. Wilson Bull. 112:165-186.
Taylor, P. B. 1998. Rails: A guide to the rails, crakes, gallinules and coots of the world. Yale University Press, New Haven, Connecticut.
Worthy, T. H., and R. N. Holdaway. 2002. The lost world of the moa. Indiana University Press, Bloomington.
Wragg, G. M. 1995. The fossil birds of Henderson Island, Pitcairn Group: Natural turnover and human impact, a synopsis. Biol. J. Linn. Soc. 56:405-414.
Jeremy J. Kirchman2,4 and David W. Steadman3
2 Department of Zoology, University of Florida, Gainesville, Florida 32611.
3 Florida Museum of Natural History, University of Florida, Gainesville, Florida 32611.
4 Corresponding author: e-mail: jkirchman@zoo .ufl.edu; phone: 352-392-1721 x232; fax: 352-392-3704.
Copyright University Press of Hawaii Apr 2006
Source: Pacific Science
www.redorbit.com/news/science/453924/new_species_of_rails_aves_rallidae_from_an_archaeological_site/index.html