Expunging the ‘Mascarene starling’ Necropsar leguati: archives, morphology and molecules topple a myth by Storrs L. Olson, Robert C. Fleischer, Clemency T. Fisher & Eldredge Bermingham
Received 8 November 2003
Until relatively recently, species of birds known from unique specimens tended to be ignored, often being written off as dubious in one way or another. Because of the rapidity of extinction on islands, however, it is not at all unlikely that a species could be collected and prepared as a museum specimen only once and then become extinct, leaving little or no other trace of its former existence. Careful study of the history of unique specimens and intensive scrutiny of the specimens themselves often leads to a species being fully restored, which has contributed significantly to our knowledge of biodiversity, zoogeography, and evolution (e.g. Graves & Olson 1987, James et al. 1989, Olson 1986a,b, Olson et al. 1989). Less frequently, investigations of an unique holotype do not end in such positive
results and specimens may turn out to be composites (Olson & Schifter 1989), hybrids (Olson & Violani 1996), or simply mislabelled and misidentified (Olson 1992). The subject of the present study, a supposed Mascarene starling, falls in the
last category. This unfortunate relic was trebucheted into ornithological legend by Henry Ogg Forbes, director of the Liverpool Museum (Forbes 1898), in a six-page paper with a colour plate (Fig. 1), line drawing and table. At a time when ornithologists were routinely introducing new species of birds with a descriptions of only a sentence or two, such detail as Forbes advanced might now seem exemplary were it not for the fact that ultimately the entire result is seen to be a fiction.
This single specimen became the focus of one unfounded assumption after another, compounded by some over-vigorous imaginations and lack of any careful subsequent scientific scrutiny, which led to the development of what in hindsight
can only be regarded as a myth. With the true identity of this specimen now at hand, its entire written history can be summed up as a banquet of codswallop.
Figure 1. The first published illustration of Necropsar leguati, from Forbes (1898)
History
The specimen in question (Liverpool Museum D.1792; Figs. 2–3) was among a mixed lot purchased 10 August 1850 from the dealer Jules Verreaux by the 13th Earl of Derby, whose collection was transferred to the Liverpool Museum in 1851
(Fisher 2002). Its identity was still undetermined when the specimen, then a flat skin, surfaced in an inventory of the collection in 1897. The bird, which was essentially white and had been labelled only ‘Madagascar’ by the Maison Verreaux,
had been placed among bulbuls of the genus Hypsipetes (Forbes 1898: 30), one Madagascan species of which is about the same size and of somewhat the same build. Thus the initial tentative placement of the specimen was among the bulbuls, Pycnonotidae. ‘That it had been left undetermined for so long a period is probably due to the fact of its being taken for an albino of some species of the above-named genus, or perhaps for a white starling’ (Forbes 1898: 30). The last phrase is a non
sequitur—if it had been stored among bulbuls, what grounds would there be for assuming that anyone had previously taken it for a starling? Forbes had evidently reached the conclusion beforehand that the specimen was a starling, and that it had come from the Mascarene islands, so his presentation was
weighted heavily towards making the few available facts fit with his preconception. Madagascar is close to the Mascarenes, and the Mascarenes were known to have been the home of two species of starlings. One of these, Fregilupus varius, is known
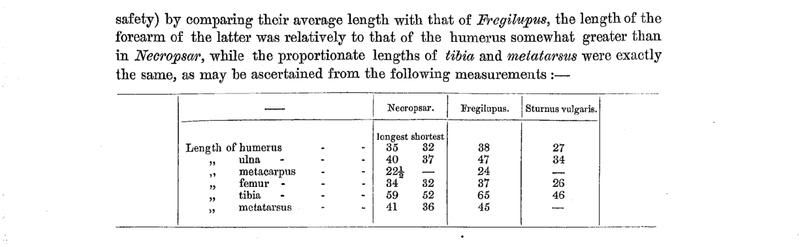
historically from the island of Réunion, where it became extinct in the middle of the 19th century (Fuller 2001). This was a showy bird, with large crest and white head and underparts, thus seemingly fitting with the whiteness of Forbes’ specimen. The other Mascarene starling is known from fossils from the island of Rodrigues and was named Necropsar rodericanus (H. H. Slater in Günther & Newton 1879: 427). Forbes referred to an anonymous early account, first published by Newton (1875), concerning a bird with white and black plumage, the size of a Blackbird Turdus merula, that had been encountered on Islet au Mât (now called I^le Gombrani) but not on the main island where the bones of Necropsar rodericanus were later found. This account was later attributed to one Tafforet, whose visit to
Rodrigues took place in 1725 (Dupon, 1973, Cheke 1987). I^le Gombrani is one of a number of small islets within the Rodrigues lagoon that would have provided refugia for the native birds that had been decimated by rats on the main island by 1725. Rats arrived as early as September 1601 during the visit of a Dutch fleet. All the islets are too small to have harboured endemic species and consequently Rodrigues would have had only one species of starling. Despite the fact that it had no black in the wings and tail, Forbes assumed that the mystery skin was the same species as in Tafforet’s anonymous account, and that this had to be different from Fregilupus varius because of its lack of a crest, among other things, and from Necropsar rodericanus because of the smaller tarsus. He
therefore named the specimen from the Derby collection as a new species, Necropsar leguati, evidently not wishing to create a new genus for it.
Rothschild (1907: 6), disliking the idea of two starlings existing on Rodrigues, suggested that N. leguati was probably an albinistic example of ‘the Mauritius species of Necropsar’. Mauritius is the only Mascarene island where no starling is
otherwise known, and there is not yet a shred of evidence, historical or paleontological, that it ever harboured a starling. Hachisuka (1953), never one to let the lack of evidence stand in the way of naming new taxa, seized upon Rothschild’s conjecture
and proceeded to create a new genus Orphanopsar, with N. leguati as its type and only species, although the only difference from N. rodericanus that he mentioned was smaller size. Earlier Hachisuka (1937) had gone even further out of bounds in placing the bird observed on Islet au Mât in the Corvidae and naming it as a new genus and species of aberrant chough, Testudophaga bicolor, based only on Tafforet’s description. This, however, has no relevance to the history of the type of N. leguati. Greenway (1958) was very circumspect and conservative, suggesting that N. leguati might be a specimen of N. rodericanus, and he listed both in the synonymy
of Fregilupus varius. Wagstaffe (1978) ventured the opinion that N. leguati was a valid species. In the influential Peters Check-list, Amadon (1962: 103) unquestioningly accepted Necropsar leguati as a valid species, giving its provenance as ‘Met [sic] Islet, off Rodriguez’, and suggesting that it was ‘possibly identical with N. rodericanus Sclater’. This was the received wisdom at least as late as 1987 (Fuller 1987).
The first hint that things might not be as they seemed came from P. J. Morgan, Wagstaffe’s successor as Keeper of Vertebrate Zoology at the Liverpool Museum (in Cheke 1987: 49), whose examination of the specimen of N. leguati suggested that it ‘may not be a starling at all.’ This proves to be the case.
Plumage and morphology As described by Forbes (1898: 34) the specimen of Necropsar leguati is ‘white everywhere, except for a lighter or darker ferruginous wash on the external webs of
the distal half of the primaries and secondaries, as also on the outer webs of the newly moulted, and on both webs of the unmoulted rectrices.’ This was repeated verbatim by Hachisuka (1953: 204), as usual without attribution or quotes. In our
examination of the specimen, however, it was obvious that the brownish portions of the remiges were asymmetrical from one side of the specimen to the other. Thus we agree completely with Morgan (in Cheke 1987: 49) that the darker portions of the
flight feathers are due to ‘discolouration’. This staining was evidently an ongoing process, because, as noted by Forbes, the older feathers of the tail are more heavily coloured than the freshly moulted ones, suggesting the bird may have been kept in
captivity. In any case, the plumage is entirely white which, with the yellowish bill and feet, which were probably pink in life, leave no doubt that the specimen is an albino. As further evidence of this, the bases of the contour feathers are white,
whereas they are dark, for example, in Fregilupus varius and Cinclocerthia.
Figure 2. X-radiographs of the skulls in skin specimens in lateral view. A, holotype of Necropsar leguati LIV D.1792; B, Cinclocerthia gutturalis BMNH 1856.3.12.10 (Martinique); C, Cinclocerthia tremula pavida BMNH 1840.5.13.11 (Nevis); D, Cinclocerthia macrorhyncha BMNH 1894.12.24.2 (St Lucia);
E, Cinclocerthia ruficauda tenebrosa BMNH 1898.2.8.21 (St Vincent).
Note not only the great similarity of N. leguati and Cinclocerthia, but also the absence of the ossified posterior border of the nostril that characterises the Pycnonotidae, with which N. leguati was first associated.
Figure 3. Specimens in ventral view: A, Cinclocerthia tremula pavida BMNH 1840.5.13.11 (Nevis); B, C. ruficauda tenebrosa BMNH 1898.2.8.21 (St Vincent); C, C. c. macrorhyncha BMNH 1894.12.24.2 (St Lucia); D, holotype of Necropsar leguati LIV D.1792.
But an albino what? Surely it is not a starling. ‘Its wings, however, are quite unlike those of any starling,’ as Forbes (1898: 30) himself stated outright. He went on to amplify this further (p. 33): ‘It possesses ten primaries, and of these the tenth, or outermost, is not the rudimentary or very reduced quill seen in the Sturnidae generally.’ It is testimony to his willingness to delude himself that he could continue to rationalise that the bird was somehow nevertheless a starling.
The well-developed outer primary not only establishes that the specimen is not a starling, but also not a member of any of the so-called nine-primaried oscines. Likewise, the wing is not ‘short, rounded, and ‘concave’ as attributed to most of the
species of Timaliidae (Newton 1896: 963). The tarsal scutellation (inaccurately depicted by Forbes 1898) is not ‘booted’ as in the typical thrushes (Turdidae). Bill
shape alone eliminates many other groups such as finches, shrikes and flycatching birds. Bulbuls (Pycnonotidae) may be absolutely ruled out by their tiny feet, bill shape and presence of filoplumes in the nape. Furthermore, X-radiographs of the
holotype of N. leguati (Fig. 2) show that it did not have the posterior margin of the nostril ossified as in the Pycnonotidae (Olson 1990), thus eliminating the family with which the specimen had originally been associated while in Lord Derby’s collection. Through such a process of elimination we were finally led to make detailed comparisons with the species of Mimidae (mockingbirds and thrashers) and here the resemblances became much greater. In size, length and shape of bill, wing
formula, and foot structure and scutellation, the holotype of N. leguati could be
Figure 4. Close-up of ventral view of head (A, B) and lateral view (C) of entire specimen of A, C, holotype of Necropsar leguati LIV D.1792; B, Cinclocerthia macrorhyncha BMNH 1894.12.24.2 (St Lucia).
matched only by the West Indian thrashers known as tremblers (Cinclocerthia) (Figs. 3–4). The tremblers range throughout most of the Lesser Antilles, from Saba to St Vincent, with six recognised taxa that have usually been considered subspecies of
Brown Trembler C. ruficauda (e.g. Davis & Miller 1960). However, in a more recent detailed analysis, Storer (1989) advocated recognition of two species, C. gutturalis for the populations from Martinique and St Lucia, and C. ruficauda for
the remainder. Studies of DNA suggest that recognition of even more species may be warranted (see below). The fact that the holotype of N. leguati came from the French establishment of Maison Verreaux suggests the possibility of the specimen having originated in the French islands of Martinique or Guadeloupe. Colour comparisons, of course, are impossible because the specimen is an albino. The various taxa differ among
themselves to some extent in size, but our measurements of the specimen of N. leguati compared with those in Storer (1989) were inconclusive, partly because there were so few specimens available from the French islands, especially Martinique. So only molecular analysis remained for a finer resolution of the
identity of N. leguati. Provenance As noted, the specimen of Necropsar leguati was received from Jules Verreaux and
is labelled only ‘Madagascar’. Forbes (1898: 30) suggested that because Verreaux had been to the Mascarene islands and had collected a specimen of the Réunion Starling Fregilupus varius that he had prepared as a skeleton, that he ‘may have also
secured’ the bird in question during that visit. But then Forbes went on to remark that ‘It is well known that M. Verreaux was often very inexact in the precise geographical data he inscribed on the labels of his specimen.’ In fact, the unreliability of specimen labels emanating from Maison Verreaux is notorious in ornithology. The firm more than once sent out material completely mislabelled’ and engaged in deliberately altering the identifications of birds’ eggs to increase their market value
(Mearns & Mearns 1988: 406). If it is allowed, as Forbes did, that the specimen in question could have come from somewhere other than Madagascar, then there is no basis for picking one locality over another without some sort of corroborating
evidence. We decided to look through some of the other specimens in the Liverpool
Museum that came from Verreaux in the same purchase lot as the holotype of N. leguati, to see if any of them might shed some light on its provenance (this action being made easier by the existence of a specimen database which could be crossreferenced by date of purchase). We were not disappointed in our expectation of finding specimens with obviously erroneous locality information: a specimen of the
North American Virginia Rail Rallus limicola, with ‘Martinique’ crossed out by Verreaux and ‘Nle Zelande’ (New Zealand) substituted; a Ptilinopus fruit dove labelled ‘Iles Marquises’ annotated by David Holyoak as having an erroneous locality; a specimen of one of the South American subspecies of Southern Roughwinged Swallow Stelgidopteryx ruficollis labelled ‘Jamaique’; etc. But the real clincher came when examining a specimen identified as Foudia madagascariensis and labelled ‘Madagascar’. Foudia is a genus of weaver finch
(Ploceidae) confined to the south-west Indian Ocean islands. The supposed Foudia D.3185) is a partial albino with the belly, mantle, rump and one rectrix white. The characteristic bill shape, rufous undertail-coverts and coloration of the remaining
pigmented parts, however, establish beyond doubt that this is a specimen of the Lesser Antillean Bullfinch Loxigilla noctis.
The presence of another albinistic bird endemic to the Lesser Antilles labelled ‘Madagascar’ in the same purchase lot as the holotype of Necropsar leguati is too unlikely to be attributed to coincidence. Verreaux was obviously receiving specimens from the Lesser Antilles from someone, probably an aviculturist, with an affinity for albinos. To these Verreaux proceeded to give what may have been considered a more interesting and exotic provenance. Mitochondrial DNA
Our comparisons were greatly facilitated by the fact that mitochondrial DNA adenosine triphosphatase subunits 8 and 6 (ATP6/8) had already been sequenced for most relevant taxa of Antillean Mimidae (Hunt et al. 2001). Grey Trembler Cinclocerthia gutturalis of Martinique was not included in that study, however, for lack of fresh material. Therefore, we obtained samples from a skin specimen of C. gutturalis (USNM 75724) and from the holotype of Necropsar leguati. We also
sequenced part of the mitochondrial cytochrome b gene (Cytb) from the same samples of Cinclocerthia and Margarops (sensu lato) as used in Hunt et al. (2001). in April 2000, a small piece of toe pad was sliced with a sterile scalpel from the
specimen of Necropsar leguati at the Liverpool Museum (D.1792). The sample was placed in a sterile microfuge tube and returned to the ancient DNA laboratory in the Department of Biological Sciences at the University of Durham, where all subsequent molecular analyses of the sample were completed. In February 2001, a small piece of toe pad was sliced with a sterile scalpel from a specimen of Cinclocerthia gutturalis (USNM 75724 male, collected by F. A. Ober on Martinique). The sample was placed in a sterile microfuge tube and returned to the ancient DNA laboratory in the National Zoological Park, Smithsonian Institution, Washington DC, where all subsequent molecular analyses of the sample were completed. Thus, DNA isolations of the two toe pad slices were conducted in separate
laboratories on separate continents. Part of each sample was cut into smaller pieces using a sterile scalpel. DNA was isolated from the diced samples using an overnight DTT / SDS / Proteinase K digestion, followed by phenol-chloroform extraction and centrifugal dialysis concentration (Fleischer et al. 2000). From 2–4 ml of DNA extract were used in PCR amplifications of specific mtDNA regions for the museum specimen samples. We targetted two mtDNA regions: Cytb and ATP8/6. A total of 332 bp of the Cytb region was amplified in two small pieces with primers Cytb1-anc and Cytb.X (112 bp of sequence) and Cytb2.RC and Cytb.wow (220 bp) (Dumbacher et al. 2003). Part of the ATP8/6 region was also amplified in two
pieces totaling 350 bp with primers A6MNH and t-lys (192 bp; Dumbacher et al. 2003) and ATP6R (5’ AGGTGTCCTGCTGTGAGGTT-3’) and ATP6L (5’- CCTAGCCTTCCCCCTATGAC-3’) (158 bp). Additional Cinclocerthia and possible outgroup ATP8/6 sequences were obtained from Genbank, from a deposit by Hunt et al. (2001). Additional Cytb sequences were obtained using Cytb1-anc
and Cytb.wow on a subsample of the fresh material from Hunt et al. (2001). PCR products were cycle sequenced and sized on an ABI automated DNA sequencer. Resulting sequence chromatograms were inspected to ensure correct nucleotide
identification. Both strands were sequenced. Sequences were aligned in Sequencher (version 4.1.2; GeneCodes Corporation) to each other and to outgroup sequences. We obtained a total of 682 bp of mtDNA sequence from Necropsar leguati and
the Martinique specimen of Cinclocerthia gutturalis: 332 of Cytb, 350 of ATP6/8 (Genbank accession numbers pending). The sequences were aligned with each other. The Cytb sequences were identical except for a few ambiguous bases in the
Figure 5. A phylogram of one of two most parsimonious trees found using a branch-and-bound search. The tree length is 135 steps. The numbers at each node are the percentage of 1,000 bootstrap replicates in which the particular node was found, if above 50%. Tree is based on 648 bp of Cytb and ATP8/6
sequence of two museum specimens (Martinique—C. gutturalis and Necropsar leguati) and Cytb sequences from this paper and ATP8,6 sequences from Hunt et al. (2001). specimen of C. gutturalis. The ATP8 and 6 sequences were identical between the two specimens, but for a single nucleotide substitution. This low level of sequence divergence strongly indicates that the two specimens are from the same species. We aligned the two museum specimen sequences to the ATP6/8 sequences from Hunt et al. (2001) and to the Cytb sequences. We conducted four different types of phylogenetic analyses on the dataset to
reconstruct phylogenetic trees from the sequences: parsimony, neighbour-joining (using a Kimura 2-parameter distance), and maximum likelihood using PAUP* 4.0b10 (Swofford 2002), and a Bayesian analysis using Mr. Bayes 3.0 (Huelsenbeck & Ronquist 2001). We show here one maximum parsimony tree with associated bootstrap values on the nodes (Fig. 5), but note that in all trees produced by any criterion, the specimen of Necropsar leguati is a clear and close sister lineage to the
specimen of C. gutturalis from Martinique. The clade is basal to a C. ruficauda/C. gutturalis clade, and this is nested within a clade of other Caribbean mimids, including Scaly-breasted Thrasher Margarops fuscus and Pearly-eyed Thrasher M.
fuscatus. Thus, based on both the sequence comparison and the phylogenetic analysis it appears that the holotype of Necropsar leguati is an example of C. gutturalis sensu stricto, which is known historically only from Martinique. On the basis of the DNA analysis, one could justify recognising as many as four
species of Cinclocerthia: C. tremula (Lafresnaye 1843) from Guadeloupe, with a subspecies C. t. pavida Ridgway (1904, type locality St Kitts) from most of the Lesser Antilles north of Guadeloupe; C. ruficauda (Gould 1836) of Dominica, with
a subspecies C. r. tenebrosa Ridgway (1904) from St Vincent; C. gutturalis (Lafresnaye 1843) from Martinique; and C. macrorhyncha Sclater (1866) from St Lucia. The distribution of the two basal taxa, C. gutturalis and C. macrorhyncha, scattered among the seemingly more derived forms, does not match expectation based on linear colonisation of the Lesser Antilles and suggests a rather old split. Such a conclusion requires corroboration from other sources such as nuclear DNA,
morphology, and perhaps call and behaviour. Regardless, Necropsar leguati clearly belongs with Cinclocerthia gutturalis.
Conclusions On the basis of morphology and the history of the specimen, the holotype of Necropsar leguati Forbes, 1898, a supposed Mascarene starling, is shown instead to be a mislabelled albino West Indian trembler of the genus Cinclocerthia. Studies of mitochondrial DNA identify the specimen with the distinctive basal taxon from Martinque C. gutturalis. These conclusions have the following consequences:
1. Necropsar leguati Forbes, 1898, becomes a junior subjective synonym of Ramphocinclus gutturalis Lafresnaye, 1843 = Cinclocerthia gutturalis.
2. Orphanopsar Hachisuka, 1953, becomes a junior subjective synonym of Cinclocerthia Gray, 1840.
3. Necropsar leguati must now be entirely expunged from the avifauna of the Mascarene islands.
www.nmnh.si.edu/vert/birds/pdf/slo3.pdf