|
|
Post by another specialist on Jun 6, 2005 15:05:34 GMT
Orthiospiza howarthi
Olson & James 1991
Holocene of Maui, e Hawaiian Islands
Primary materials: Holotype: partial associated skeleton
Secondary materials: Paratypes: maxillae, mandibulae, humerus
Storrs L. Olson & Helen F. James,
Descriptions of thirty-two new species of birds from the Hawaiian Islands: Part II. Passeriformes
Ornithological Monographs 46 (1991)
The American Ornithologists' Union, Washington D.C.
|
|
|
|
Post by another specialist on Nov 8, 2005 7:39:00 GMT
|
|
|
|
Post by another specialist on Nov 8, 2005 7:39:21 GMT
commonly called the Highland Finch
|
|
|
|
Post by another specialist on Jul 22, 2006 9:20:36 GMT
Orthiospiza, new genus
Type species: Orthiospiza howarthL new species.
Included species: Type-species only.
Distribution: Maui: higher elevations on the south and east slopes of Mr. Haleakala.
Etymology: Greek, orthios, high, lofty, and spiza, a finch, to emphasize that
specimens of this genus have so far been found only at relatively high elevations.
The gender is feminine.
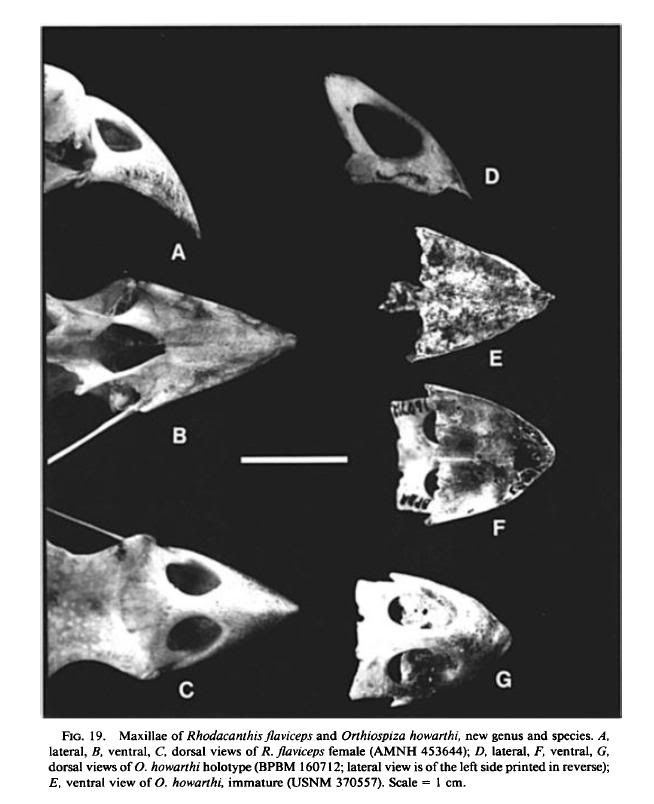
Diagnosis: Large drepanidines with arched, finch-like bills, distinguished by the
greatly enlarged narial openings, the absence of a ventral trough on the maxilla,
and the straight rather than indented posteroventral contour of the maxilla. The
cranial fenestra is smaller than in any other finch-billed drepanidine in which the
cranium is known, except Chloridops kona. The muscle scars on the cranium and
mandible reflect a well-developed musculature for adduction of the mandible,
with robust zygoraatic processes, prominent interorbital cristae, a distinct crest
delimiting the main area of attachment of adductor muscles on the skull, and
partly ossified aponeuroses preserved on the coronoid processes and on the ventral
margins of the mandibular foramina in the adult specimen. By contrast, the
palatines are underdeveloped, with the transpalatine processes short and weak
and the prepalatine bars very weak compared to Rhodacanthis and other large
finches. The prepalatine bars also differ from the relatively thin bars of Psittirostra
in being very short rather than elongated. The surfaces on the maxilla for articulation
of the palatines and jugals are distinctly weaker than expected for a finch
with such a sturdy bill.
The maxilla is broad and pointed, with an arched dorsal surface. The tomial
crest of the maxilla is evenly decurved, as in Telespiza, and the dorsal nasal bar
is elongated, as in Psittirostra. Broad, moderately excavated lateral grooves provide
the only relief on the otherwise nearly flat ventral surface of the maxilla. The
ventral ridges are very close-set, as in Chloridops kona, but extend nearly to the
tip of the maxilla, as in Telespiza and Loxioides. The ventral ridges differ from
other genera in being not at all prominent, and in being rugose in the adult.
Between these ridges, where the medial trough usually occurs, the maxilla is flat
and is also rugose in the adult.
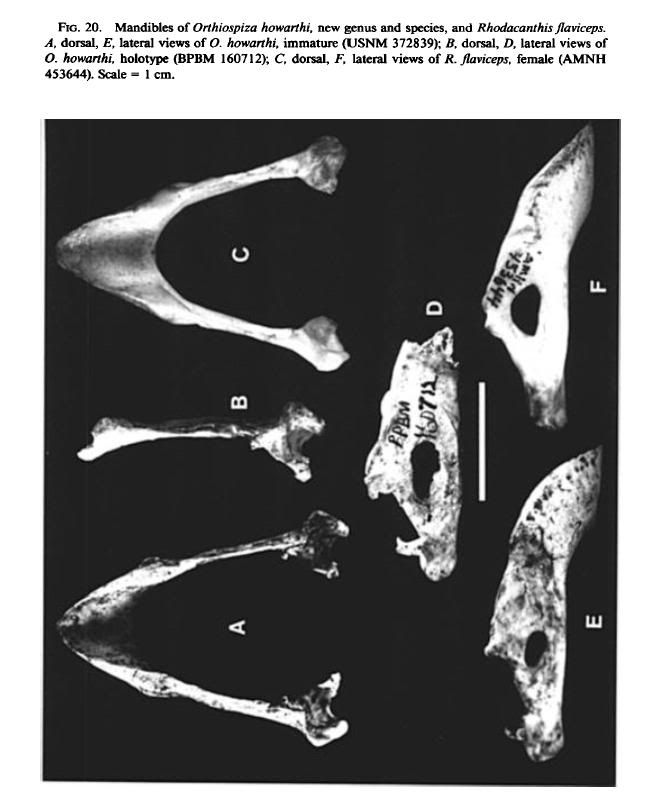
The mandible is deep and pointed as in Rhodacanthis, but has a short symphysis
and a strongly and evenly decurved tomial crest as in Loxioides. The shape of
the lateral cotylae, which are elevated, angled posteromedially, and have the
anterior portion not much wider than the posterior portion, is reminiscent of the
much smaller cotylae of Xestospiza fastigialis, new genus and species. |
|
|
|
Post by another specialist on Jul 22, 2006 9:22:03 GMT
|
|
|
|
Post by another specialist on Jul 22, 2006 9:23:56 GMT
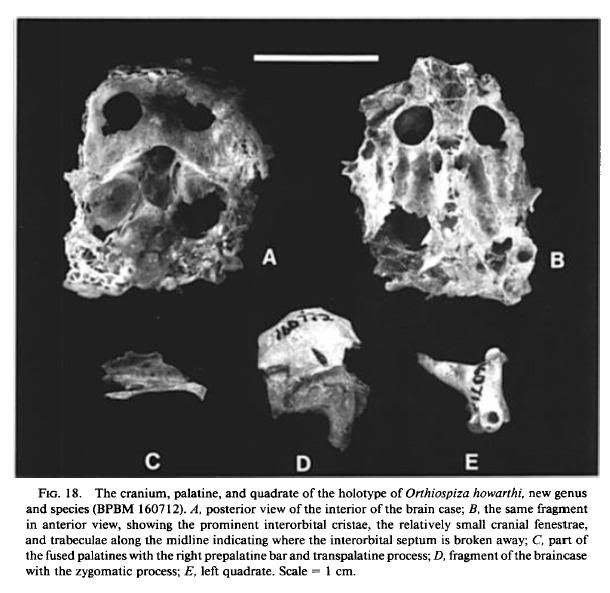
Orthlosl)i•a howarthi, new species
(Figs. 18, 19D-G, 20A-B, D-E, 21)
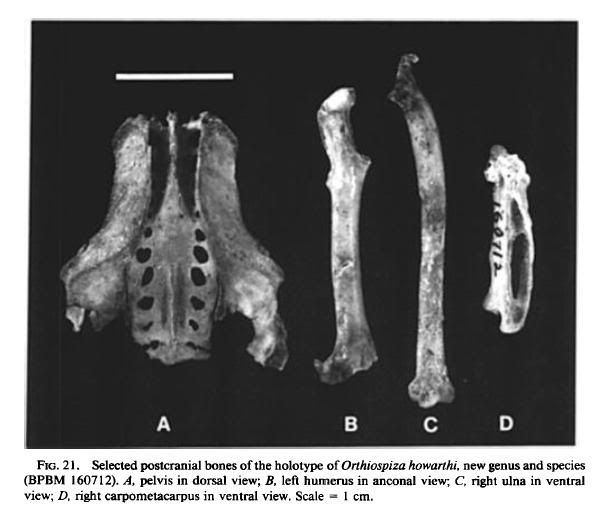
Holotype: Partial associated skeleton, BPBM 160712. Collected between 24
February and 4 March 1984 by Francis G. Howarth, Fred D. Stone, Betsy H.
Gagng and others. The specimen consists of the maxilla lacking the tip (Fig. 19D,
F, G), fragments of the cranium preserving the posterior wall of the orbits (Fig.
18A, B), the basitemporal plate, the left zygomatic process (Fig. 18D), and the
supraoccipital, the right pterygoid, the fused palatines with the left side mostly
missing (Fig. 18C), both quadrates (Fig. 18E), both rami of the mandible but not
the symphysis (Fig. 20B, D), the axis and eight additional cervical vertebrae, three
thoracic vertebrae, two caudal vertebrae, the pelvis (Fig. 21A), the anterior half
of the fight scapula, the dorsal half of the fight coracoid, the ventral third of the
left coracoid, both humeri (each missing parts of both ends; Fig. 21B), the right
ulna (Fig. 21C), fragments of the shaft of the left ulna, both carpometacarpi with
damaged proximal ends (Fig. 21D), both femora with damaged distal ends, the
right tibiotarsus with damaged ends, the broken left tibiotarsus, proximal ends
with parts of the shafts of both tarsometatarsi, the damaged distal end of the left
tarsometatarsus, three pedal phalanges, and some unidentified bone fragments.
Type locality: Lua Manu (1,830 m), Kipahulu Valley, Maul, Hawaiian Islands.
The location of the holotype within the cave coresponds with "Site L" on the
cave map drawn by Fred Stone and Francis Howarth, a copy of which is on file
with the accession record at the Bishop Museum (BPBM Accession 1984.116).
Distribution: Maul: Lua Manu, Puu Makua Cave, and Auwahi Cave.
Etymology: To Francis G. Howarth, for his studies of the entomology of Hawaiian
caves and for his attentivehess to potential fossil deposits in lava tubes.
Measurements (ram) of holotype: The comparable measurement of Rhodacanthis
palmeri (AMNH 453623) is given in parentheses when available. Humerus:
length (estimated), 25.0 (24.5); proximal width, 7.2 (7.8); mid-shaft width, 2.4
(2.4). Ulna: length, 29.8. Carpometacarpus: length, 16.2. Synsacrum: length, 2.5.
Pelvis: width between antitrochanters, 19.1. Femur: length, 26.9; proximal width,
4.8; mid-shaft depth, 1.9. Tibiotarsus: length without cnemial crest, 41.5. Tarsometatarsus:
proximal width, 4.3 (4.2); mid-shaft width, 1.7 (1.7). For additional
measurements of holotype, see Tables 7 and 8.
Paratypes: Auwahi Cave: Maxilla lacking lateral nasal bars, USNM 370557
(Fig. 19E); complete mandible, USNM 372839 (Fig. 20A, E); humerus lacking
the head and distal condyles, USNM 372840. These bones appear to be from a
single juvenile individual.
Puu Makua Cave: Complete maxilla, not fully mature, USNM 445797; mandible
in two pieces, lacking the anterior two-thirds of the right ramus, not fully
mature, USNM 445798. These bones may be from a single individual.
Measurements ofparatypes: See Tables 7 and 8.
Diagnosis: As for the genus.
Remarks: The body size is comparable to Rhodacanthis palmeri, the largest of
the finch-billed drepanidines for which post-cranial bones are available to compare.
The relative completeness of the holotype of O. howarthi permits us to make
some additional osteological comparisons beyond those mentioned in the generic
diagnosis. The fragments of cranium preserve evidence of the drepanidine affinities
of Orthiospiza: the interorbital septurn has widely separated walls enclosing a
network of bony trabeculae; the posterolateral margin of the cranial fenestra is
straight; the anterior wall of the cranial cavity between the cranial fenestrae is
broad and flat, not blade-like. Together these conditions define a synapomorphous
character that occurs only in the Carduelinae, including the Drepanidini (Zusi
1978). The palatine process of the premaxilla is fused to the prepalatine bar in
the configuration of a lateral flange, another derived character that is present in
cardueline finches, but also appears in other, unrelated passerines (Book 1960).
Orthiospiza has many presumably primitive characters in common with the
other finch-billed drepanidine genera. The palatines have a very broad blade,
short transpalatine processes and prepalatine bars, and have the walls of the
palatine hasp wide-set to embrace the thick basisphenoid rostrum. The quadrate
has the typical stocky form found in finch-billed drepanidines, with a thick body
and a rounded medial condyle set on a short neck. The large narial opening in
Orthiospiza retains the subcircular shape that is typical of heavy-billed finches,
rather than being elongated anteroposteriorly as is true of the relatively weakbilled
Psittirostra and Melarnprosops, and of all thin-billed drepanindines except
Hernignathus wilsoni and H. lucidus. The broad pelvis is also characteristic of the
finch-billed as opposed to the thin-billed drepanidines.
Autapomorphous characters that distinguish Orthiospiza from other drepanidines
are the greatly enlarged but not elongated narial openings, the absence of
a medial trough on the maxilla, and the straight posteroventral edge of the maxilla.
Orthiospiza shares at least one character with each of several other finch-billed
genera, although none of these stands out as convincing evidence of relationship:
the reduced size of the cranial fenestra is shared with Chloridops, the weak lateral
nasal bars and pointed bill are shared with Rhodacanthis, and the weak ventral
ridges that extend nearly to the tip of the maxilla resemble Loxioides and Telespiza.
Two of the three known specimens of Orthiospiza are juveniles. These were
not found in owl pellet deposits but appear to be remains of birds that entered
the caves unaided. A high proportion of juveniles dying in caves might be a
reflection of species-specific behaviors such as cavity nesting or exploratory foraging
by young birds.
Orthiospiza is one of relatively few avian fossil taxa that may have been restricted
to montane habitats in the Hawaiian Islands. All of the specimens originate
from three lava tubes located at relatively high elevations (1,145, 1,463, and 1,830
m). No specimens have yet been found in Puu Naio Cave (305 m), the most
productive site for fossil passefines on Maui so far.
It is a striking circumstance that the only two passefine skeletons preserved in
the Upper Kipahulu Valley caves are of an extinct thrush and O. howarthi, while
the abundant native species that occur in the vicinity now are not represented in
the cave. |
|
|
|
Post by another specialist on Jul 22, 2006 9:29:04 GMT
|
|
|
|
Post by another specialist on Jul 22, 2006 9:29:22 GMT
|
|
|
|
Post by another specialist on Jul 22, 2006 9:29:51 GMT
|
|